T019 · Molecular dynamics simulation¶
Note: This talktorial is a part of TeachOpenCADD, a platform that aims to teach domain-specific skills and to provide pipeline templates as starting points for research projects.
Authors:
Pietro Gerletti, CADD seminar 2020, Charité/FU Berlin
Mareike Leja, 2020/21, Internship at Volkamer Lab, Charité
Jeffrey R Wagner, 2020, Open Force Field Consortium
David Schaller, 2020/21, Volkamer Lab, Charité
Andrea Volkamer, 2020/21, Volkamer Lab, Charité
Note
This talktorial was designed to be used with Google Colab. It is also possible to use it on a local computer. However, performing the molecular dynamics simulation may take a considerably long time if no GPU is available.
Also, note that this talktorial will not run on Windows for the time being (check progress in this issue).
Aim of this talktorial¶
In this talktorial, we will learn why molecular dynamics (MD) simulations are important for drug design and which steps are necessary to perform an MD simulation of a protein in complex with a ligand. The kinase EGFR will serve as sample system for simulation.
Contents in Theory¶
Molecular dynamics
Force fields
Boundary conditions
MD simulations and drug design
EGFR kinase
Contents in Practical¶
Installation on Google Colab
Adjust environment for local installations running on Linux or MacOS
Import dependencies
Download PDB file
Prepare the protein ligand complex
Protein preparation
Ligand preparation
Merge protein and ligand
MD simulation set up
Force field
System
Perform the MD simulation
Download results
References¶
Review on the impact of MD simulations in drug discovery (J Med Chem (2016), 59(9), 4035‐4061)
Review on the physics behind MD simulations and best practices (Living J Comp Mol Sci (2019), 1(1), 5957)
Review on force fields (J Chem Inf Model (2018), 58(3), 565-578)
Review on EGFR in cancer (Cancers (Basel) (2017), 9(5), 52)
Summarized statistical knowledge from Pierre-Simon Laplace (Théorie Analytique des Probabilités Gauthier-Villars (1820), 3)
Inspired by a notebook form Jaime Rodríguez-Guerra (github)
Repositories of OpenMM and OpenMM Forcefields, RDKit, PyPDB, MDTraj, PDBFixer
Wikipedia articles about MD simulations, AMBER and force fields in general
[1]:
import sys
on_colab = "google.colab" in sys.modules
if on_colab:
%pip install teachopencadd --no-deps -q
!teachopencadd -d 19
%pip uninstall teachopencadd -y -q
%pip install -qr requirements.txt
%conda install mdtraj openmm openmmforcefields openff-toolkit pdbfixer pypdb rdkit -y -c conda-forge
Theory¶
Molecular dynamics¶
Molecular dynamics is a computational method analyzing the movements and interactions of atoms and molecules of a defined system. The method stems from theoretical physics, where it was developed in the 1950s (Alder and Wainwright in J Chem Phys (1959), 31(2), 459), although the ideas behind it can be dated much earlier:
An intelligence which could, at any moment, comprehend all the forces by which nature is animated and the respective positions of the beings of which it is composed, and moreover, if this intelligence were far-reaching enough to subject these data to analysis, it would encompass in that formula both the movements of the largest bodies in the universe and those of the lightest atom: to it nothing would be uncertain, and the future, as well as the past, would be present to its eyes. The human mind offers us, in the perfection which it has given to astronomy, a faint sketch of this intelligence. (Pierre-Simon Laplace, 1820)
Let us just take this statement by Laplace as the ideological substrate underneath molecular dynamics simulations. In other terms, we can approximate the behavior of a physical system by knowing the characteristics of its components and applying Newton’s laws of motion. By solving the equations of motion, we can obtain a molecular trajectory of the system, which is a series of snapshots with the positions and velocities of all its particles, as well as its potential energy. To do so, we define functions, called force fields, which provide an approximate description of all the forces applied to each particle in the system. We then use numerical integrators to solve the initial value problem for the system and obtain the trajectory. As it sounds, the process requires quite a bit of processing power and it was only few years ago that MD started seeing a more widespread use, especially in the field of computational chemistry and biology, as well as in drug discovery (J Med Chem (2016), 59(9), 4035‐4061).

Figure 1: Molecular dynamics simulation of the rotation of a supramolecule composed of three molecules in a confined nanoscopic pore (Palma et al. via Wikimedia).
Force fields¶
Force fields describe the forces between atoms within and between molecules. They are parametric equations with components for different forces (bond stretching, van-der-Waals and more). The parameter values are usually derived experimentally and change for each MD scenario, depending on the molecules involved and the simulation settings. The result is a mathematical description of the energy landscape of the system, in which the forces acting on each particle result from the gradient of the potential energy with respect to the coordinates of the atoms.
Several force fields are available, each with its own characteristics (J Chem Inf Model (2018), 58(3), 565-578). In this notebook, we will use a member of the AMBER force field family, which are widely used for MD simulations of proteins. Their functional form is:
The formula consists of a sum of different components. The first three components contain information about bond lengths, angles and torsions (intramolecular forces). The last component describes intermolecular, non-bonded forces like van-der-Waals forces and electrostatic interactions. The various parameters, denoted by a superscript 0, depend on the force field used and vary between all members of the AMBER force field family. Note that these force fields assume fixed-charge particles and do not allow polarization, nor do they consider how a local charge influences its surroundings.
The following visual representation of force fields components shows the same concepts in a more intuitive way.

Figure 2: Components of a molecular mechanics force field (Edboas via Wikimedia).
Boundary conditions¶
Often, molecular systems are simulated in a box filled with solvent such as water. These boxes are of finite size, which results in problems for molecules at or near the box boundaries. With which molecules should those interact? Periodic boundary conditions can avoid such boundary artifacts by simulating a theoretically infinite system. Molecules at one boundary of the box thereby interact with molecules at the boundary on the other side of the box. This mimics a situation, in which the simulation box is surrounded by replicas of itself. When visualizing such MD simulations, one can often observe that particles leave the box at one side (Fig. 3). However, they re-appear at the same time on the other side of the box with the same velocity. For simulations under periodic boundary conditions, it is recommended to use a simulation box large enough, so that the simulated macromolecule does not come into contact with neighboring images of itself.

Figure 3: Molecular dynamics simulation of water molecules with periodic boundary conditions (Kmckiern via Wikimedia).
{kind=link}
MD simulations and drug design¶
MD simulations give valuable insights into the highly dynamic process of ligand binding to their target. When a ligand (or a drug) approaches a macromolecule (protein) in solution, it encounters a structure in constant motion. Also, ligands may induce conformational changes in the macromolecule that can best accommodate the small molecule. Such conformations may not be discovered with static methods. Accordingly, binding sites that are not observed in static ligand-free structures, but can be discovered with MD simulations, are sometimes called cryptic binding sites (J Med Chem (2016), 59(9), 4035‐4061). The identification of such binding sites with MD simulation can kickstart new drug discovery campaigns. Later in the drug discovery process, MD simulations can also be used to estimate the quality of computationally identified small molecules before performing more costly and time-intensive in vitro tests. Altogether, MD simulations pose a valuable asset in computational drug design.
EGFR kinase¶
The Epidermal Growth Factor Receptor (EGFR) is an important drug target with implications in cancer and inflammation (Wikipedia). It is a transmembrane protein with an extracellular receptor domain and an intracellular kinase domain. The binding of the endogenous ligand epidermal growth factor results in activation of the kinase domain via dimerization and autophosphorylation. The activated kinase domain can then phosphorylate downstream signaling proteins triggering DNA synthesis and cell proliferation (Cancers (Basel) (2017), 9(5), 52). Inhibition of this kinase is the underlying mechanism of action of several already approved cancer drugs (DrugBank). In this talktorial, we use the PDB structure 3POZ of this kinase, which is in complex with the small molecule inhibitor 03P, to perform an MD simulation (PDB: 3POZ).

Figure 4: Structure 3POZ of the EGFR kinase domain bound to inhibitor 03P (PDB: 3POZ).
Practical¶
We will now proceed to perform an MD simulation using the molecular dynamics engine OpenMM, a high performance toolkit for molecular simulation. It is open source and can be used as application or library. We will use it as Python library.
Import dependencies¶
[2]:
import copy
from pathlib import Path
import requests
from IPython.display import display
import numpy as np
from rdkit import Chem
from rdkit.Chem import Draw
from rdkit.Chem import AllChem
import mdtraj as md
import pdbfixer
import openmm as mm
import openmm.app as app
from openmm import unit
from openff.toolkit.topology import Molecule, Topology
from openmmforcefields.generators import GAFFTemplateGenerator
[3]:
# create data directory if not exists
HERE = Path(_dh[-1])
DATA = HERE / "data"
DATA.mkdir(exist_ok=True)
Download PDB file¶
The Protein Data Bank (PDB) allows for easy downloading of files via url.
[4]:
pdbid = "3POZ"
ligand_name = "03P"
pdb_path = DATA / f"{pdbid}.pdb"
pdb_url = f"https://files.rcsb.org/download/{pdbid}.pdb"
[5]:
r = requests.get(pdb_url)
r.raise_for_status()
with open(pdb_path, "wb") as f:
f.write(r.content)
Prepare the protein ligand complex¶
Protein preparation¶
A crucial part for successful simulation is a correct and complete system. Crystallographic structures retrieved from the Protein Data Bank often miss atoms, mainly hydrogens, and may contain non-standard residues. In this talktorial, we will use the Python package PDBFixer to prepare the protein structure. However, co-crystallized ligands are not handled well by PDBFixer and will thus be prepared separately.
[6]:
def prepare_protein(
pdb_file, ignore_missing_residues=True, ignore_terminal_missing_residues=True, ph=7.0
):
"""
Use pdbfixer to prepare the protein from a PDB file. Hetero atoms such as ligands are
removed and non-standard residues replaced. Missing atoms to existing residues are added.
Missing residues are ignored by default, but can be included.
Parameters
----------
pdb_file: pathlib.Path or str
PDB file containing the system to simulate.
ignore_missing_residues: bool, optional
If missing residues should be ignored or built.
ignore_terminal_missing_residues: bool, optional
If missing residues at the beginning and the end of a chain should be ignored or built.
ph: float, optional
pH value used to determine protonation state of residues
Returns
-------
fixer: pdbfixer.pdbfixer.PDBFixer
Prepared protein system.
"""
fixer = pdbfixer.PDBFixer(str(pdb_file))
fixer.removeHeterogens() # co-crystallized ligands are unknown to PDBFixer
fixer.findMissingResidues() # identify missing residues, needed for identification of missing atoms
# if missing terminal residues shall be ignored, remove them from the dictionary
if ignore_terminal_missing_residues:
chains = list(fixer.topology.chains())
keys = fixer.missingResidues.keys()
for key in list(keys):
chain = chains[key[0]]
if key[1] == 0 or key[1] == len(list(chain.residues())):
del fixer.missingResidues[key]
# if all missing residues shall be ignored ignored, clear the dictionary
if ignore_missing_residues:
fixer.missingResidues = {}
fixer.findNonstandardResidues() # find non-standard residue
fixer.replaceNonstandardResidues() # replace non-standard residues with standard one
fixer.findMissingAtoms() # find missing heavy atoms
fixer.addMissingAtoms() # add missing atoms and residues
fixer.addMissingHydrogens(ph) # add missing hydrogens
return fixer
[7]:
# prepare protein and build only missing non-terminal residues
prepared_protein = prepare_protein(pdb_path, ignore_missing_residues=False)
Prepare ligand¶
After preparing the protein, we turn our attention to the ligand. Again, we need to add hydrogens, but also need to make sure the bond orders are correctly assigned, since some PDB entries may contain errors. We use the Python package RDKit, an open source cheminformatics library. We will provide the correct protonation state and bond orders to RDKit via a SMILES string. Uncharged isomeric SMILES strings for each co-crystallized ligand can be found in their respective PDB entry. The ligand of PDB entry 3POZ has the name 03P. If a ligand is likely to bind in its charged form or as a specific tautomer, such characteristics need to be incorporated into the SMILES string.
[8]:
def prepare_ligand(pdb_file, resname, smiles, depict=True):
"""
Prepare a ligand from a PDB file via adding hydrogens and assigning bond orders. A depiction
of the ligand before and after preparation is rendered in 2D to allow an inspection of the
results. Huge thanks to @j-wags for the suggestion.
Parameters
----------
pdb_file: pathlib.PosixPath
PDB file containing the ligand of interest.
resname: str
Three character residue name of the ligand.
smiles : str
SMILES string of the ligand informing about correct protonation and bond orders.
depict: bool, optional
show a 2D representation of the ligand
Returns
-------
prepared_ligand: rdkit.Chem.rdchem.Mol
Prepared ligand.
"""
# split molecule
rdkit_mol = Chem.MolFromPDBFile(str(pdb_file))
rdkit_mol_split = Chem.rdmolops.SplitMolByPDBResidues(rdkit_mol)
# extract the ligand and remove any already present hydrogens
ligand = rdkit_mol_split[resname]
ligand = Chem.RemoveHs(ligand)
# assign bond orders from template
reference_mol = Chem.MolFromSmiles(smiles)
prepared_ligand = AllChem.AssignBondOrdersFromTemplate(reference_mol, ligand)
prepared_ligand.AddConformer(ligand.GetConformer(0))
# protonate ligand
prepared_ligand = Chem.rdmolops.AddHs(prepared_ligand, addCoords=True)
prepared_ligand = Chem.MolFromMolBlock(Chem.MolToMolBlock(prepared_ligand))
# 2D depiction
if depict:
ligand_2d = copy.deepcopy(ligand)
prepared_ligand_2d = copy.deepcopy(prepared_ligand)
AllChem.Compute2DCoords(ligand_2d)
AllChem.Compute2DCoords(prepared_ligand_2d)
display(
Draw.MolsToGridImage(
[ligand_2d, prepared_ligand_2d], molsPerRow=2, legends=["original", "prepared"]
)
)
# return ligand
return prepared_ligand
Calling this function with the isomeric SMILES string taken from the PDB entry for 03P returns a correctly prepared ligand. Printed is a 2D-representation of the original and the prepared ligand for inspection.
[9]:
smiles = "CC(C)(O)CC(=O)NCCn1ccc2ncnc(Nc3ccc(Oc4cccc(c4)C(F)(F)F)c(Cl)c3)c12"
rdkit_ligand = prepare_ligand(pdb_path, ligand_name, smiles)
[10:10:57] WARNING: More than one matching pattern found - picking one
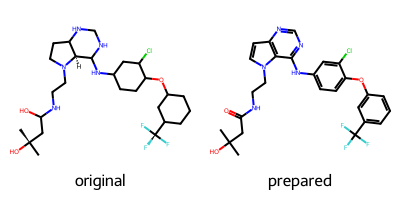
Merge protein and ligand¶
In the next step, we want to merge the prepared protein and ligand structures using the Python package MDTraj. MDTraj can handle the prepared protein, which is currently a PDBFixer molecule, a format that has a topology and atom positions similar to and usually interchangeable with OpenMM Modeller topologies and positions. For the ligand however, we need to do several conversions, since it is currently an RDKit molecule.
[10]:
def rdkit_to_openmm(rdkit_mol, name="LIG"):
"""
Convert an RDKit molecule to an OpenMM molecule.
Inspired by @hannahbrucemcdonald and @glass-w.
Parameters
----------
rdkit_mol: rdkit.Chem.rdchem.Mol
RDKit molecule to convert.
name: str
Molecule name.
Returns
-------
omm_molecule: openmm.app.Modeller
OpenMM modeller object holding the molecule of interest.
"""
# convert RDKit to OpenFF
off_mol = Molecule.from_rdkit(rdkit_mol)
# add name for molecule
off_mol.name = name
# add names for atoms
element_counter_dict = {}
for off_atom, rdkit_atom in zip(off_mol.atoms, rdkit_mol.GetAtoms()):
element = rdkit_atom.GetSymbol()
if element in element_counter_dict.keys():
element_counter_dict[element] += 1
else:
element_counter_dict[element] = 1
off_atom.name = element + str(element_counter_dict[element])
# convert from OpenFF to OpenMM
off_mol_topology = off_mol.to_topology()
mol_topology = off_mol_topology.to_openmm()
mol_positions = off_mol.conformers[0]
# convert units from Ångström to nanometers
# since OpenMM works in nm
mol_positions = mol_positions.to("nanometers")
# combine topology and positions in modeller object
omm_mol = app.Modeller(mol_topology, mol_positions)
return omm_mol
[11]:
omm_ligand = rdkit_to_openmm(rdkit_ligand, ligand_name)
Now protein and ligand are both in OpenMM like formats and can be merged with MDTraj.
[12]:
def merge_protein_and_ligand(protein, ligand):
"""
Merge two OpenMM objects.
Parameters
----------
protein: pdbfixer.pdbfixer.PDBFixer
Protein to merge.
ligand: openmm.app.Modeller
Ligand to merge.
Returns
-------
complex_topology: openmm.app.topology.Topology
The merged topology.
complex_positions: openmm.unit.quantity.Quantity
The merged positions.
"""
# combine topologies
md_protein_topology = md.Topology.from_openmm(protein.topology) # using mdtraj for protein top
md_ligand_topology = md.Topology.from_openmm(ligand.topology) # using mdtraj for ligand top
md_complex_topology = md_protein_topology.join(md_ligand_topology) # add them together
complex_topology = md_complex_topology.to_openmm()
# combine positions
total_atoms = len(protein.positions) + len(ligand.positions)
# create an array for storing all atom positions as tupels containing a value and a unit
# called OpenMM Quantities
complex_positions = unit.Quantity(np.zeros([total_atoms, 3]), unit=unit.nanometers)
complex_positions[: len(protein.positions)] = protein.positions # add protein positions
complex_positions[len(protein.positions) :] = ligand.positions # add ligand positions
return complex_topology, complex_positions
[13]:
complex_topology, complex_positions = merge_protein_and_ligand(prepared_protein, omm_ligand)
/home/michael/.teachopencadd_envs/teachopencadd_T019_py312/lib/python3.12/site-packages/openmm/unit/quantity.py:750: UnitStrippedWarning: The unit of the quantity is stripped when downcasting to ndarray.
self._value[key] = value / self.unit
[14]:
print("Complex topology has", complex_topology.getNumAtoms(), "atoms.")
# NBVAL_CHECK_OUTPUT
Complex topology has 5561 atoms.
MD simulation set up¶
We can now use the prepared complex to set up the MD simulation.
Force field¶
Common force fields like AMBER have parameters for amino acids, nucleic acids, water and ions and usually offer several options to choose from depending on your aim. We use the amber14-all.xml force field file, which is shipped with OpenMM and includes parameters for proteins, DNA, RNA and lipids. For solvation we use the standard three-site water model TIP3P.
Parameters for ligands however are not included. To generate these parameters, we can use the General AMBER ForceField (GAFF), which is implemented in the Python package OpenMM Forcefields. The following function generates a force field object holding standard AMBER parameters and additionally includes parameters for a small molecule if required.
[15]:
def generate_forcefield(
rdkit_mol=None, protein_ff="amber14-all.xml", solvent_ff="amber14/tip3pfb.xml"
):
"""
Generate an OpenMM Forcefield object and register a small molecule.
Parameters
----------
rdkit_mol: rdkit.Chem.rdchem.Mol
Small molecule to register in the force field.
protein_ff: string
Name of the force field.
solvent_ff: string
Name of the solvent force field.
Returns
-------
forcefield: openmm.app.Forcefield
Forcefield with registered small molecule.
"""
forcefield = app.ForceField(protein_ff, solvent_ff)
if rdkit_mol is not None:
gaff = GAFFTemplateGenerator(
forcefield="gaff-2.2.20",
molecules=Molecule.from_rdkit(rdkit_mol, allow_undefined_stereo=True),
)
forcefield.registerTemplateGenerator(gaff.generator)
return forcefield
[16]:
forcefield = generate_forcefield(rdkit_ligand)
System¶
With our configured force field we can now use the OpenMM Modeller class to create the MD environment, a simulation box which contains the complex and is filled with a solvent. The standard solvent is water with a specified amount of ions. The size of the box can be determined in various ways. We define it with a padding, which results in a cubic box with dimensions dependent on the largest dimension of the complex.
Note this step can take a long time, in the order of minutes, depending on your hardware.
[17]:
modeller = app.Modeller(complex_topology, complex_positions)
modeller.addSolvent(forcefield, padding=1.0 * unit.nanometers, ionicStrength=0.15 * unit.molar)
With our solvated system and force field, we can finally create an OpenMM System and set up the simulation. Additionally to the system the simulation needs an integrator. An OpenMM Integrator defines a method for simulating a system by integrating the equations of motion. The chosen Langevin Integrator uses Langevin equations. A list of all different kinds of integrators can be found in the OpenMM Docs. For further insight into the Langevin Integrator, we recommend reading about Langevin equations, e.g. on Wikipedia.
[18]:
system = forcefield.createSystem(modeller.topology, nonbondedMethod=app.PME)
integrator = mm.LangevinIntegrator(
300 * unit.kelvin, 1.0 / unit.picoseconds, 2.0 * unit.femtoseconds
)
simulation = app.Simulation(modeller.topology, system, integrator)
simulation.context.setPositions(modeller.positions)
Perform the MD simulation¶
Now that everything is set up, we can perform the simulation. We need to set starting positions and minimize the energy of the system to get a low energy starting configuration, which is important to decrease the chance of simulation failures due to severe atom clashes. The energy minimized system is saved.
[19]:
simulation.minimizeEnergy()
with open(DATA / "topology.pdb", "w") as pdb_file:
app.PDBFile.writeFile(
simulation.topology,
simulation.context.getState(getPositions=True, enforcePeriodicBox=True).getPositions(),
file=pdb_file,
keepIds=True,
)
Once the minimization has finished, we can perform the MD simulation. In this talktorial, we will do a short simulation for illustration. Simulations for research purposes span several nanoseconds, even up to microseconds. We will simulate only 100 ps of molecular dynamics corresponding to 50k steps of 2 fs each. We save molecular “snapshots” every 10 ps (5000 steps), for a total of 10 frames. The results are saved in an .xtc file, which contains the coordinates of all the atoms at a given time point. Together with the PDB file of the energy minimized system written before, it gives us all the information needed for later analysis.
Note: This talktorial will only generate a 20 fs trajectory, if not on Google Colab. However, if you have a good GPU available, you can also increase the simulation time.
[20]:
# output settings
if on_colab:
steps = 50000 # corresponds to 100 ps
write_interval = 5000 # write every 10 ps
log_interval = 2500 # log progress to stdout every 5 ps
else:
steps = 10 # corresponds to 20 fs
write_interval = 1 # write every 2 fs
log_interval = 1 # log progress to stdout every 2 fs
simulation.reporters.append(
md.reporters.XTCReporter(file=str(DATA / "trajectory.xtc"), reportInterval=write_interval)
)
simulation.reporters.append(
app.StateDataReporter(
sys.stdout,
log_interval,
step=True,
potentialEnergy=True,
temperature=True,
progress=True,
remainingTime=True,
speed=True,
totalSteps=steps,
separator="\t",
)
)
The velocities for all particles in the system are randomly chosen from a distribution at the given temperature. We chose 300 Kelvin, which is some degrees above room temperature. A random seed is generated, but could be explicitly given to reproduce results.
Then the simulation is performed by taking the steps defined before.
[21]:
simulation.context.setVelocitiesToTemperature(300 * unit.kelvin)
simulation.step(steps) # perform the simulation
#"Progress (%)" "Step" "Potential Energy (kJ/mole)" "Temperature (K)" "Speed (ns/day)" "Time Remaining"
10.0% 1 -911672.8833984158 300.58994432283663 0 --
20.0% 2 -898806.4210639504 267.2506776332451 1.78 0:00
30.0% 3 -883111.4814176012 238.60290950665717 1.73 0:00
40.0% 4 -867596.4630740668 196.76725273444427 1.68 0:00
50.0% 5 -862553.2471993582 175.87229216182052 1.6 0:00
60.0% 6 -854763.8296435549 163.39398429953755 1.54 0:00
70.0% 7 -852074.9736099411 147.9384677561452 1.5 0:00
80.0% 8 -855085.7518770137 152.24611569256365 1.5 0:00
90.0% 9 -856760.1494785415 157.91401106119835 1.49 0:00
100.0% 10 -861616.5007841537 165.39716192526913 1.49 0:00
[22]:
# Check the trajectory exists and is not empty
(DATA / "trajectory.xtc").stat().st_size > 0
# NBVAL_CHECK_OUTPUT
[22]:
True
Download results¶
You can execute the following cell if you are working on Google Colab to download the MD simulation results.
[23]:
if on_colab:
files.download(DATA / "topology.pdb")
files.download(DATA / "trajectory.xtc")
Discussion¶
Quiz¶
Which inter- and intramolecular forces are being considered in the AMBER force field? Can you think of any forces not taken into account?
Would you expect to see the exact same simulation results when running the notebook twice with the same parameters?
Try doing a short (10ps, snapshot every 1ps) simulation of a protein without a ligand. You can find a broad variety of structures on PDB or you can use the EGFR kinase and remove the ligand.
Further reading¶
Enhanced sampling methods¶
In theory, unbiased MD simulations should be capable of simulating binding and unbinding events of a drug molecule and its macromolecular target. However, the timescale of binding and unbinding events lies in the millisecond to second range. Enhanced sampling methods aim to accelerate the conformational sampling (J Med Chem. 2016, 59(9), 4035-61).
One of these is Free energy perturbation (FEP) (also called alchemical free energy calculation), which computes the free energy difference when going from a state A to another state B. It is often employed in lead optimization to evaluate small modification at the ligand, that may boost the binding affinity for the desired target. The ligand from state A is thereby gradually transformed into the ligand of state B by simulating several intermediate (“alchemical”) states (alchemistry).
Another technique for free-energy calculations is Umbrella sampling (US). US enforces sampling along a collective variable (CV) by performing staged simulations with an energetic bias. The bias usually takes the form of a harmonic potential, hence the term “umbrella”. Its goal is to sample high-energy regions along the CV. However, the use in drug design is limited by the high computational cost.
In contrast, Steered MD (SMD) follows a different approach: it applies external forces to the system. Those forces are time-dependent and facilitate the unbinding of the ligand from the target. The SMD calculates the final force exerted on the system. The unbinding force profile can then be used to filter hits from docking calculations and to discriminate active from inactive molecules.